|
http://www.duiops.net/seresvivos/metazoos_desa.html
El desarrollo embrionario
El desarrollo embrionario es el período desde la fecundación hasta el nacimiento del nuevo ser, aunque no exista fecundación, como sucede en los casos de partenogénesis.
Consta de las fases de: fecundación, segmentación, gastrulación y organogénesis.
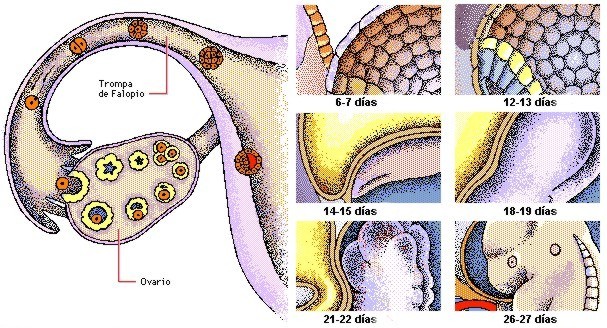
Fecundación: es la unión de las dos células reproductoras, de s.exos contrarios, los gametos, hasta que se funden en uno solo los respectivos núcleos y parte del citoplasma. Es un proceso complicado que conduce a la formación de una célula, el cigoto o huevo y que comienza con la penetración de un espermatozoide en un óvulo. En la fecundación no participa todo el espermatozoide, sino sólo el núcleo y el centrosoma; ambos corpúsculos se dirigen al núcleo femenino y el primero acaba por fusionarse con él, mientras el centrosoma se divide en dos, originándose las esferas atractivas, que se colocan en los polos del cigoto para la primera división del desarrollo embrionario, que comienza con la segmentación.
Segmentación: es la repetida división por mitosis del óvulo fecundado hasta llegar al estado de blástula, dando lugar a numerosos blastómeros. Puede ser, según la participación de todo el vitelo o la distinción en formativo y nutritivo, total o parcial; la primera puede ser igual o desigual, y la segunda discoidal o superficial. En esta fase de distinguen las siguientes formaciones:
Blastómeros: son cada una de las células en que se divide el huevo o cigoto para dar lugar a las primeras fases embrionarias.
Mórula: es el estado temprano del desarrollo de un huevo fecundado, durante el período de segmentación, en el que el conjunto de células, en número reducido todavía, se semeja a una mora. Los blastómeros emigran hacia la periferia para formar una única capa.
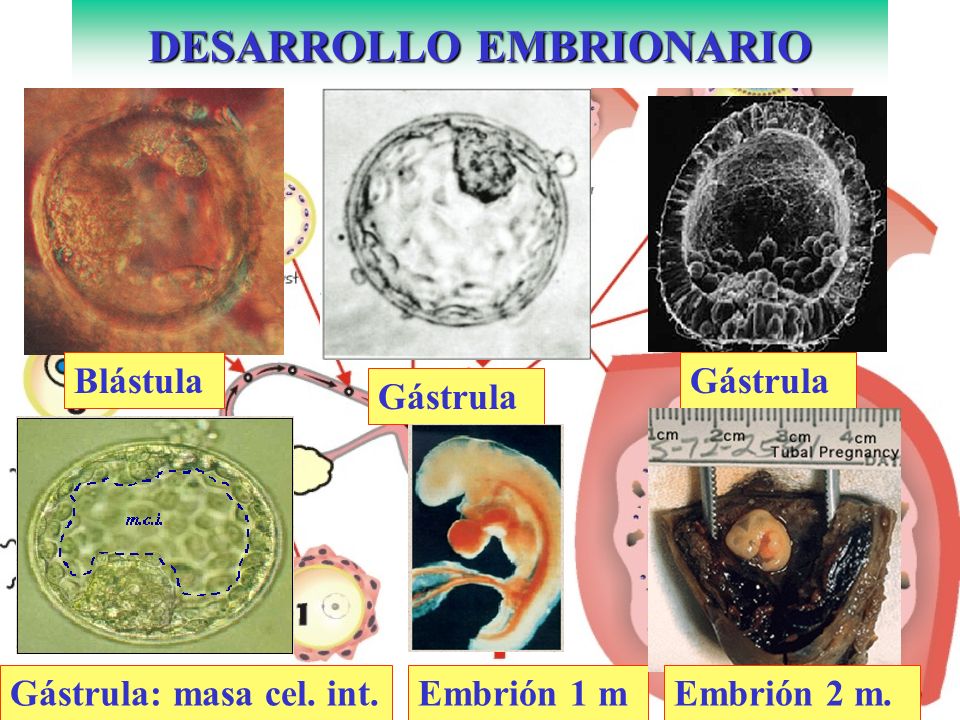
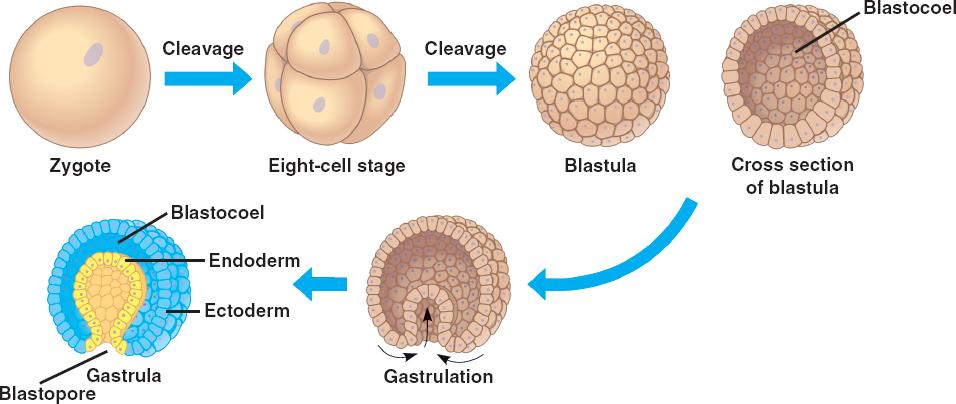
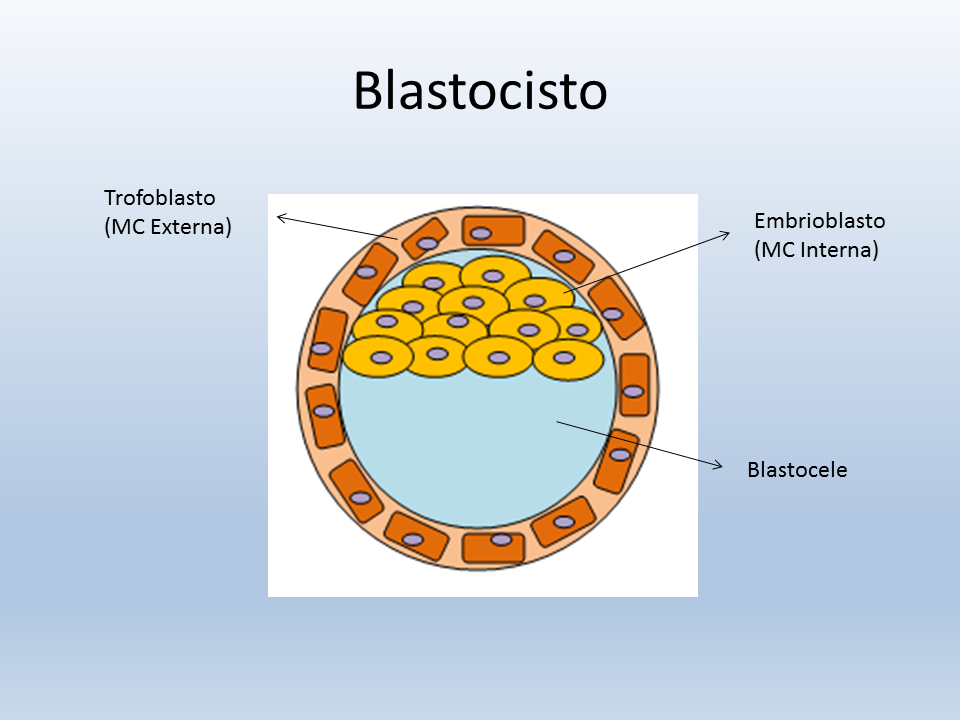
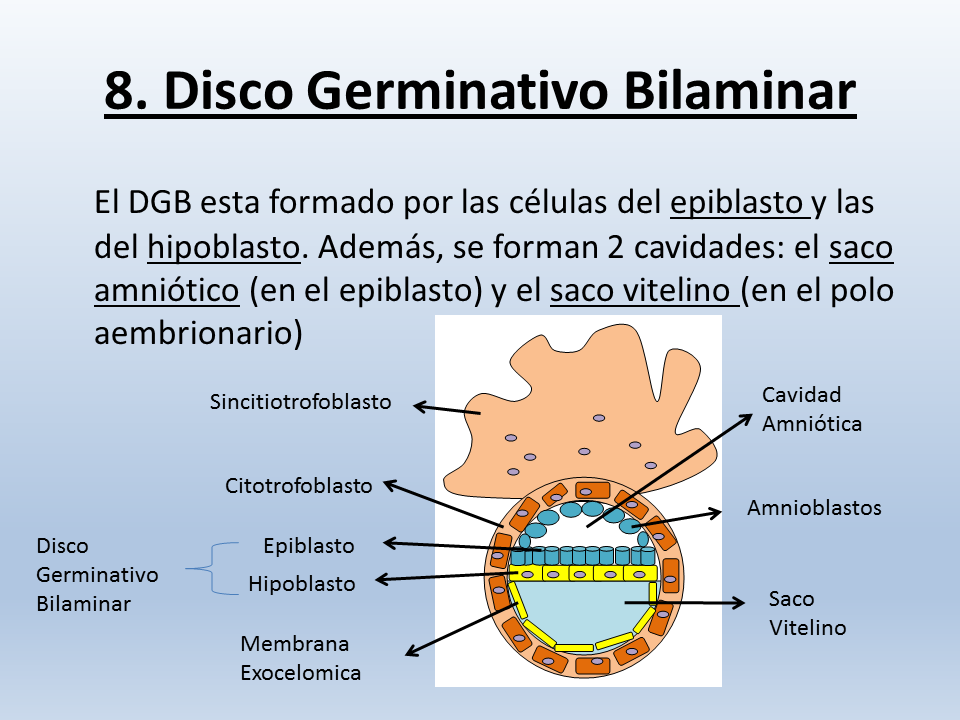
Blástula: es una de las primeras fases del desarrollo embrionario de los animales metazoos; la que sique a la mórula. Los blastómeros se disponen en una capa celular continua que circunda una cavidad interior, el blastocele, también llamada cavidad de segmentación. Sus paredes luego estarán cerradas por el blastodermo, que son los blastómeros que, dispuestos en una sola capa, forman la pared de la blástula y maracan el final de la segmentación. El blastocele está lleno de un líquido, el blastoquilo.
La estructura de la blástula es, pues, mododérmica, y su forma, muy variada, depende de la cantidad de vitelo contenida en el huevo. Por un proceso de invaginación se transforma en gástrula. El vitelo es el protoplasma del óvelo de los animales y, por extensión, del óvulo fecundado. Se distingue un vitelo germinativo, también llamado formativo o activo, que es el que experimenta la división se segmentación embrionaria, y un vitelo nutritivo o pasivo, constituido por substancias de reserva, para nutrir al embrión en las primeras fases de su desarrollo.
En los óvulos de los mamíferos vivíparos, como en los de numerosos invertebrados, predomina el vitelo germinativo en los óvulos alecitos y heterolecitos; en los de los artrópodos es variable la cantidad de vitelo nutritivo en los óvulos centrolecitos; mientras que éste ocupa la casi totalidad de la yema en los óvulos telolecitos de aves y reptiles. Algunos biólogos dan el nombre de vitelo al nutritivo o deutoplasma.
Gastrulación: es el proceso de formación de la gástrula. Comprende la invaginación o embolia, que es la forma ordinaria de la grastrulación de la blástula, consistente en que una parte de la misma se introduce en la otra, como sucede cuando se comprime una pelota de goma pinchada hasta formar con ella un casquete hemisférico: la parte que queda fuera viene a ser el ectodermo de la gástrula, y la parte invaginada el endodermo.
La gástrula es una fase del desarrollo embrionario de los metazoos, que sucede a la de blástula, y que produce en general por invaginación de ésta, con formación de un saco de pared doble, cuya cavidad, el intestino primitivo, arquenterón, celenterón o gastrocele, comunica con el exterior por un orificio, el blastoporo, que actúa de boca y a.no. Las dos capas parietales o blastodérmicas son el extodermo, la externa y el endodermo, la interna, aquél procedente de las células del polo animal de la blástula y éste de las del polo vegetativo.
Algunos animales, como los celentéreos, terminan su desarrollo en esta fase, carecen, por tanto, de cavidad general o celoma, que es una cavidad o cavidades mesodérmicas, y son los acelomados diploblásticos, con sólo dos capas blastodérmicas.
En otros aparece una tercera capa o mesodermo, producida por el ectodermo y el endodermo, en los metazoos triploblásticos, y en la mayoría de éstos, desde los briozoos a los cordados, después de haber aparecido unos esbozos mesodérmicos, se forma una cavidad general o celoma, después de haberse escindido en dos capas, la esplácnica, que junto con el endodermo de los metazoarios superiores forma el tubo digestivo, y la somática, que, unida al ectodermo, constituye la pared del cuerpo, dejando entre ambas dicha cavidad, el celoma. En los anélidos, equinodermos y cordados, el celoma es la cavidad donde se halla el tubo digestivo, mientras que en los artrópodos y moluscos se halla reducido a las cavidades de las gónadas y del aparato excretor.
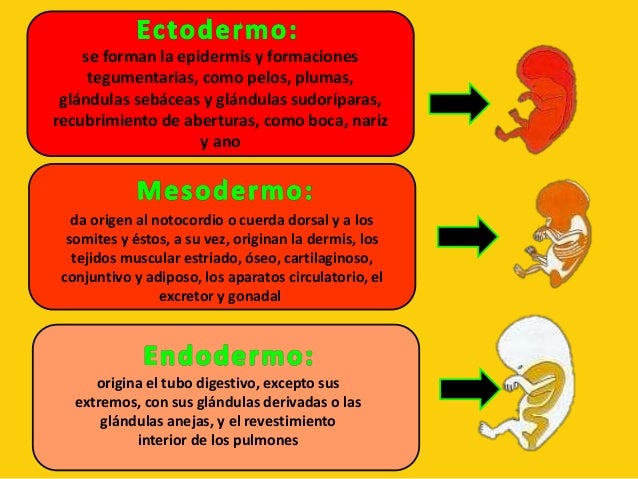
A partir del ectodermo se forman la epidermis y formaciones tegumentarias, como pelos, plumas, glándulas sebáceas y glándulas sudoríparas, recubrimiento de aberturas, como boca, nariz y a.no, los dos extremos del tubo digestivo, el sistema nervioso central y nervios periféricos, los tegumentos (que son la epidermis y sus derivados), el sistema nervioso, determinadas partes de los órganos sensoriales y las porciones extremas del tubo digestivo; el mesodermo da origen al notocordio o cuerda dorsal y a los somites y éstos, a su vez, originan la dermis, los tejidos muscular estriado, óseo, cartilaginoso, conjuntivo y adiposo, los aparatos circulatorio, el excretor y gonadal y las pleuras; el endodermo origina el tubo digestivo, excepto sus extremos, con sus glándulas derivadas o las glándulas anejas, y el revestimiento interior de los pulmones. El blastoporo gastrular se convierte en a.no en los celomados llamados deuteróstomos, equinodermos y cordados; y pasa a ser boca en los próstomos, que son todos los demás filos.
En el extremo opuesto al blastoporo, según el grupo zoológico al que pertenezca, aparece otra abertura, que actuará como boca en los deuteróstomos y como boca en los próstomos.
Organogénesis: Es la formación de los esbozos organógenos y diferenciación de los mismos.
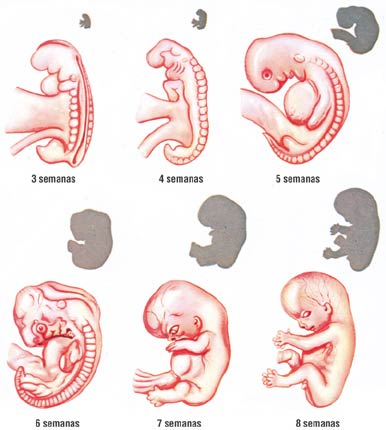
Terminado el desarrollo embrionario, el animal surge al exterior, bien por la eclosión del huevo, como en los reptiles y aves, bien en el acto del parto, como en la inmensa mayoría de los mamíferos. En numerosos peces y anfibios, como en muchos invertebrados, parte del desarrollo embrionario se realiza en la vida libre, y se continua insensiblemente, con las metamorfosis que conducen al estado adulto.
Hay dos hechos embriogénicos de gran interés:
Que la segmentación del huevo y procesos posteriores varían según la constitución de aquél.
Que en los estados embrionarios de grupos de origen común, se acentúa la semejanza de los mismos en relación con los que ofrecen los adultos; así por ejemplo, todos los embriones de los vertebrados presentan branquias, que desaparecen durante el proceso embrionario, excepto en los peces y en algunos anfibios.
Responder
|

 a cortisona, esto aísla al feto del cortisol materno. En la medida que el feto madura, la actividad 11β-HSD II aumenta. Sin embargo, hacia el final del embarazo, el hígado y los pulmones expresan actividad 11-cetoesteroide reductasa que promueve la conversión local de la cortisona en cortisol. La adrenal fetal secreta aldosterona en bajas cantidades en la mitad del embarazo, posteriormente aumenta y persiste durante el primer año de vida extrauterina. El eje hipotálamo-hipófisis-adrenales (HHA) fetal se activa progresivamente en la gestación tardía y con ello incrementa la biosíntesis de hormona liberadora de corticotropina (CRH) y arginina vasopresina (AVP) en el hipotálamo, la secreción de ACTH en la hipófisis y la sensibilidad de las enzimas esteroidogénicas a la ACTH en la corteza suprarrenal. La ACTH es secretada por las células corticotropas de la hipófisis en respuesta a la CRH y la AVP. Los estrógenos placentarios, a través de un asa de retroalimentación positiva, incrementan progresivamente la actividad del eje HHA fetal, lo cual resulta en aumento de ACTH y cortisol en las últimas 10 semanas del embarazo. Esto ayuda a la maduración visceral y pulmonar. La placenta también libera CRH y ACTH en la circulación fetal para estimular el eje HHA fetal. La CRH también interviene en el inicio del trabajo de parto. Los glucocorticoides y mineralocorticoides adrenales actúan a través de dos receptores nucleares, GR y MR. Los GR están presentes desde la mitad del embarazo en placenta, pulmón, cerebro, hígado e intestino. Los MR están presentes en los tejidos fetales a partir de la 12ª-16ª semana.
a cortisona, esto aísla al feto del cortisol materno. En la medida que el feto madura, la actividad 11β-HSD II aumenta. Sin embargo, hacia el final del embarazo, el hígado y los pulmones expresan actividad 11-cetoesteroide reductasa que promueve la conversión local de la cortisona en cortisol. La adrenal fetal secreta aldosterona en bajas cantidades en la mitad del embarazo, posteriormente aumenta y persiste durante el primer año de vida extrauterina. El eje hipotálamo-hipófisis-adrenales (HHA) fetal se activa progresivamente en la gestación tardía y con ello incrementa la biosíntesis de hormona liberadora de corticotropina (CRH) y arginina vasopresina (AVP) en el hipotálamo, la secreción de ACTH en la hipófisis y la sensibilidad de las enzimas esteroidogénicas a la ACTH en la corteza suprarrenal. La ACTH es secretada por las células corticotropas de la hipófisis en respuesta a la CRH y la AVP. Los estrógenos placentarios, a través de un asa de retroalimentación positiva, incrementan progresivamente la actividad del eje HHA fetal, lo cual resulta en aumento de ACTH y cortisol en las últimas 10 semanas del embarazo. Esto ayuda a la maduración visceral y pulmonar. La placenta también libera CRH y ACTH en la circulación fetal para estimular el eje HHA fetal. La CRH también interviene en el inicio del trabajo de parto. Los glucocorticoides y mineralocorticoides adrenales actúan a través de dos receptores nucleares, GR y MR. Los GR están presentes desde la mitad del embarazo en placenta, pulmón, cerebro, hígado e intestino. Los MR están presentes en los tejidos fetales a partir de la 12ª-16ª semana.